Tackling common tansy in North America
Common tansy is an aromatic Eurasian plant species with a long history of use as a medicinal herb. Introduced for this purpose to North America, it has since become invasive. One reason for this could be the absence of the natural enemies that keep it in check in its area of origin. CABI has been tasked with identifying specialist natural enemies from Eurasia that can be introduced into North America as biological control agents.











Project Overview
So, what’s the problem
Common tansy (Tanacetum vulgare) is a Eurasian plant species whose wide native distribution ranges from western Europe eastwards to Japan. It was introduced to North America for medicinal purposes in the 17th century. It has since become an invasive weed and is declared noxious in several US states and Canadian provinces. Common tansy is unpalatable to cattle and horses and therefore especially problematic in pastures and hayfields where it significantly reduces fodder production.
What is this project doing?
In 2006, a consortium of Canadian and American funders was established and CABI’s weed team at our centre in Switzerland was asked to start a biological control programme.
A thorough survey of the published literature revealed 169 insect species and one mite that attack Tanacetum spp. in Eurasia. Of those, 30 species only attack plants in the Tanacetum genus. We prioritised a dozen candidate insects, based on their potential host range (the plants they can develop on) and damage to the target weed.
We carried out surveys across Europe to find and collect these species. We then established rearing colonies and tests to confirm their host specificity (a guiding principle for biological control is a biological control agent should not damage plants other than the target weed).
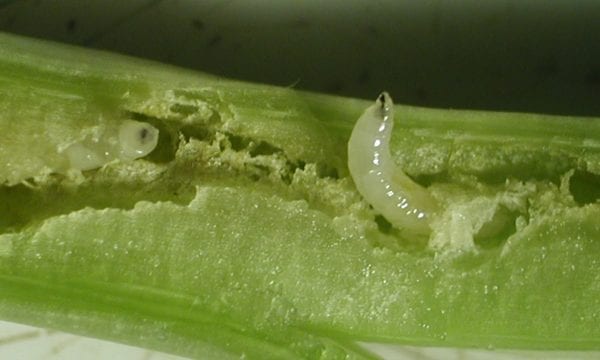
We have done this for eight potential biological control agents: the stem-boring cerambycid beetle Phytoecia nigricornis, the leaf-feeding chrysomelid beetles Longitarsus noricus, Cassida stigmatica and Chrysolina eurina, the stem-boring weevil Microplontus millefolii, the shoot- and flower-feeding gelechiid moth Isophrictis striatella, the leaf, stem and flower gall midge Rhopalomyia tanaceticola and the shoot-boring moth Gillmeria ochrodactyla (syn. Platyptilia ochrodactyla).
Results
Host-specificity tests conducted between 2006 and 2014 with P. nigricornis, L. noricus, Cassida stigmatica, I. striatella, R. tanaceticola and Chrysolina eurina showed that they are not sufficiently specific for field release in North America.
Tests with the stem-mining weevil, M. millefolii, showed more promise. Exposing weevils first to a single test species or the target weed (no-choice testing) and then a test species together with the target weed (multiple-choice testing) revealed only three test species that occasionally supported egg laying and larval development. Since only very few larvae were found on these non-target species compared with T. vulgare, our North American collaborators submitted a petition for field release to Canadian and U.S. authorities. In Canada, a release permit was granted in 2026, and the first releases of M. millefolii took place in spring 2026.
Work with the shoot-boring moth, G. ochrodactyla, has been hampered by its complicated life cycle. Females lay their eggs in summer between the individual flowers of the compact flowerhead. The following spring, the larvae burrow short mines into newly growing shoots. Their feeding can reduce growth of young shoots by over 70%. No-choice tests (where the moth is exposed to only one test species or the target weed) in cages revealed that a few non-target species are accepted for oviposition. In open-field tests, conducted in Germany and Switzerland, larvae were only found in T. vulgare and, to a much lower level, in T. huronense. Host-specificity tests with this species are currently ongoing.
Project Manager

Sonja Stutz
Research Scientist, Weed Biological Control
Rue des Grillons 1 CH-2800 Delémont, Switzerland